
Funciones de la vista
En 1943, el oftalmólogo francés André Rochon-Duvigneaud publicó el libro Les yeux et la vision des vertébrés, donde acuñó la siguiente frase: «un ave es un ala guiada por un ojo». Sin embargo, para el ornitólogo británico Graham Martin, experto en los sentidos de las aves, es más apropiada esta frase: «un ave es un pico guiado por un ojo».
Con los conocimientos actuales, se considera que la principal función de la visión, no es el vuelo sino la alimentación. Para ello, se necesita en primer lugar localizar el alimento y después, conseguirlo utilizando el pico o las patas.




Es importante manejar el pico o las patas con exactitud para conseguir un alimento inmóvil.
Y, aún se requiere más precisión, si el alimento se mueve y puede escapar-




La precisión en el uso del pico, también es necesaria para la construcción de nidos complejos y la alimentación de los pollos, pero la reproducción, a diferencia de la alimentación, solo ocurre en determinadas épocas del año.

La segunda función de la vista es la detección de depredadores y, en tercer lugar, el vuelo.

Graham Martin pone como ejemplo a las aves que tienen mayor agudeza visual como son águilas y buitres, que suelen vivir en espacios abiertos, donde pueden detectar la comida a una distancia extraordinaria para nuestra vista y no en lugares como bosques, en los que deben sortear obstáculos con gran rapidez durante el vuelo.
Tamaño de los ojos
La importancia de la visión en las aves queda reflejada por el tamaño de los ojos. A pesar de que los ojos son estructuras pesadas, la mayoría de las aves tienen los ojos grandes en relación con su pequeña cabeza. El vertebrado terrestre con los ojos más grandes es el avestruz (Struthio camelus) que tiene un diámetro de 50 mm, mientras que el diámetro del ojo humano es de 24 mm (Martin, 1985). Sin embargo, si tenemos en cuenta la relación tamaño de los ojos y tamaño corporal, los ojos más grandes se encuentran en rapaces, tanto diurnas como nocturnas.

Tener los ojos grandes es ventajoso ya que la distancia focal es larga, permite que se forme una imagen grande en la retina y puede contener un mayor número de fotorreceptores, lo que proporciona una mayor agudeza visual.
En aves con hábitos crepusculares o nocturnos como muchas rapaces, los ojos grandes no les aportan más agudeza visual, sino una mayor sensibilidad. Una pupila grande permite que entre mayor cantidad de luz, de esta manera pueden aprovechar mejor la escasa luz de que disponen en su entorno. La pupila grande también reduce la difracción. En el caso del avestruz, el ojo más grande le aumenta la sensibilidad, no la agudeza. Se cree que una mayor sensibilidad le permite protegerse de sus depredadores durante la noche.
Párpados
Las aves tienen un párpado superior y otro inferior que, salvo excepciones (por ejemplo búhos), no utilizan para parpadear. El párpado inferior es más móvil que el de los humanos.

En el borde libre de ambos párpados se suelen encontrar un tipo de plumas llamadas cerdas a modo de pestañas.

La membrana nictitante (del latín nictari = parpadear), es un repliegue de la conjuntiva que puede ser transparente, translúcida u opaca.


Por la contracción de los músculos piramidal y cuadrado, la membrana nictitante barre la superficie del ojo, horizontal u oblicuamente con gran rapidez, por lo que es difícil de ver. De esta manera extiende las lágrimas por la superficie del ojo y también lo protege.



Mismo vídeo reproducido a menor velocidad para una mejor visualización de la membrana nictitante
En algunas especies, como la urraca (Pica pica), se puede apreciar mejor, a simple vista, la membrana nictitante.
Conjuntiva
De manera similar a los mamíferos, las aves tienen una fina membrana que tapiza la superficie interior de los párpados (conjuntiva parpebral) y la parte anterior del globo ocular (conjuntiva ocular o bulbar). La conjuntiva no recubre la córnea. La conjuntiva parpebral se continúa con la conjuntiva bulbar y entre ellas se encuentra el saco conjuntival.
En la conjuntiva se encuentra tejido linfoide que contribuye a la inmunidad del ojo.
Glándulas lagrimales
Las glándulas lagrimales se localizan en el interior de la cavidad orbitaria. En las aves se pueden encontrar dos tipos: las glándulas lagrimales pequeñas y las glándulas lagrimales mayores o glándulas de Harder, llamadas así en honor a J.J. Harder que las describió por primera vez en 1694.
Las lágrimas son una secreción mucosa que contiene agua con sales disueltas como el cloruro de sodio, proteínas y lípidos. Las lágrimas mantienen el ojo húmedo y limpio, proporcionan nutrientes a la córnea y desempeñan un importante papel inmunológico.
Las glándulas lagrimales pequeñas, las glándulas de Harder y el tejido linfoide de la conjuntiva, forman parte de lo que se conoce como tejidos linfoides asociados con la cabeza (HALT). De estos, el más importante es la glándula de Harder que proporciona la mayor cantidad de anticuerpos.
Forma del globo ocular
En 1942, Walls describió tres formas de ojo aviar:
- Forma plana = Se encuentra en la mayoría de las aves. El diámetro horizontal (distancia entre el centro de la córnea y del cristalino hasta la retina) es más corto que el diámetro vertical. Se caracteriza por proporcionar un campo de visión amplio.

- Forma globosa = El diámetro horizontal y el vertical son similares. Esta forma la presentan especies diurnas que necesitan una gran agudeza visual, como águilas, halcones, cuervos, etc.

- Forma tubular = El diámetro horizontal es mayor que en las formas anteriores por lo que sobresale del cráneo. La curvatura de la córnea es grande. Es típica de rapaces nocturnas que requieren una mayor sensibilidad a la luz.

El globo ocular
En general, la estructura del globo ocular de las aves es similar a la de los demás vertebrados. Sin embargo, presenta diferencias que han mejorado la visión hasta tal punto que, en esta clase de vertebrados, se encuentran los animales que poseen la mayor agudeza visual.
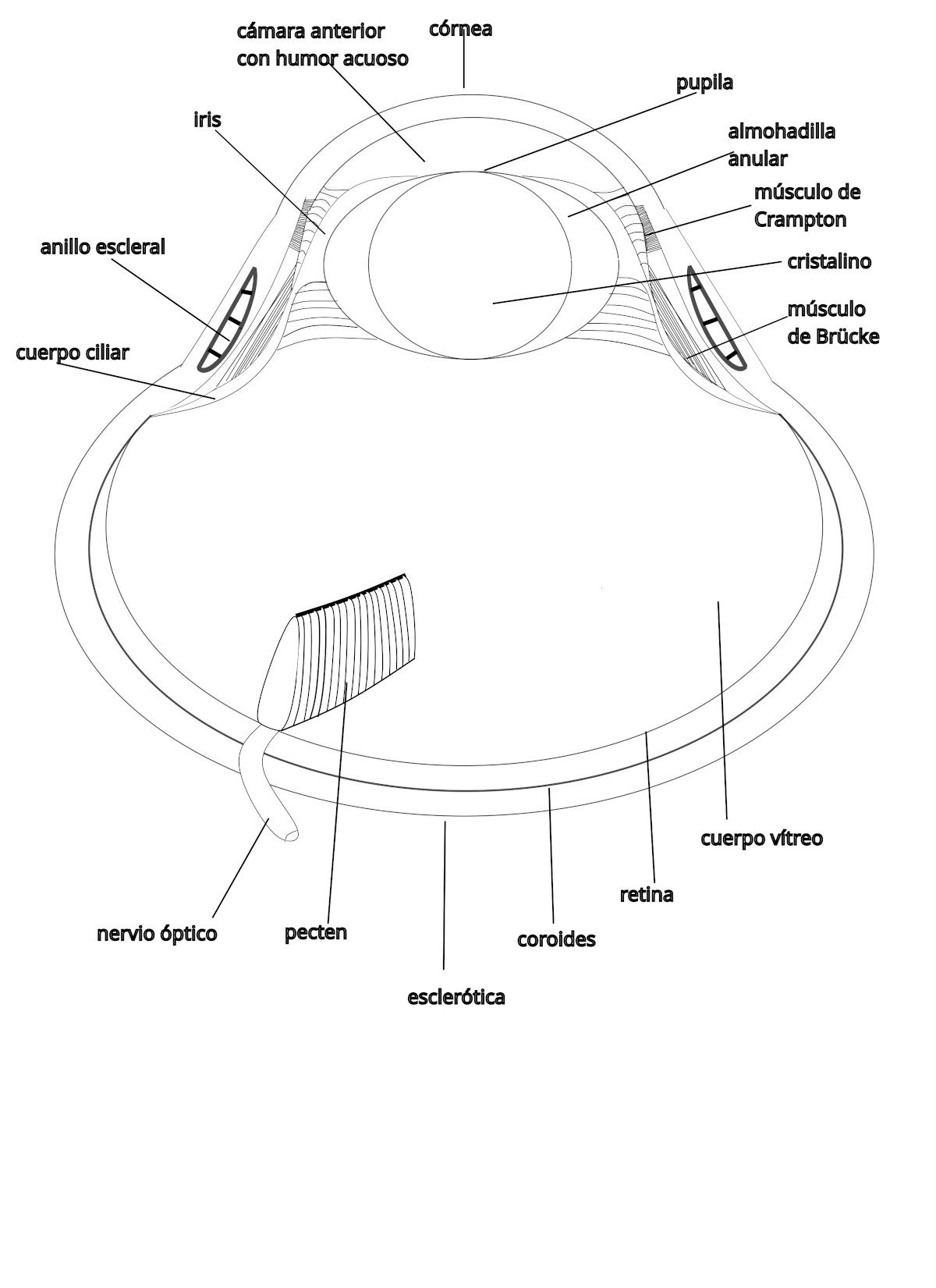
El globo ocular está formado por tres capas concéntricas: esclerótica, coroides y retina. El globo ocular de las aves no es esférico, por lo que necesita huesos y cartílago para mantener su forma.
La esclerótica es una capa fibrosa que ocupa la posición más externa. La parte anterior de la esclerótica es transparente y se llama córnea. La unión esclerocorneal es cóncava y recibe el nombre de limbo. Esta zona está reforzada por una serie de placas óseas superpuestas, los huesecillos esclerales, que se disponen formando un anillo, su número varía entre 10 y 18. El anillo escleral rodea a la córnea en su parte anterior y contribuye al mantenimiento de la forma del globo ocular.

En la región esclerocorneal se localizan los músculos de Crampton y de Brücke que intervienen en la acomodación. En el limbo se encuentra también el canal de Schlemm, seno venoso de la esclerótica, por el que drena el humor acuoso, evitando que aumente la presión intraocular. La parte posterior de globo ocular está sostenida por una lámina cartilaginosa, el cartílago escleral.
La córnea es transparente para que pueda pasar la luz y curvada.


La córnea tiene poder de refracción ya que separa dos medios con índices de refracción diferentes, como son el aire exterior y el fluido del humor acuoso. Por lo tanto, cuando la luz atraviesa la córnea cambia de velocidad y dirección.
La capa intermedia del globo ocular es la coroides. Es una capa muy vascularizada y oscura ya que contiene melanocitos. Interviene en la nutrición del globo ocular.
En la parte anterior de la coroides se encuentra el iris y el cuerpo ciliar.
El iris es una estructura musculosa en cuyo centro hay una abertura llamada pupila. La contracción de los músculos del iris aumenta o disminuye el diámetro de la pupila, de esta manera, se controla la cantidad de luz que entra en el ojo. Un ejemplo de ave que puede dilatar mucho la pupila es el pingüino rey (Aptenodytes patagonicus). Su pupila puede pasar desde los aproximadamente 0,65 mm de diámetro hasta unos 12,7 mm. Se cree que bucea en busca de alimento hasta los 200 ó 300 m de profundidad y, al abrir tanto la pupila, puede captar la escasa luz que hay incluso al mediodía (Martin, G.R. 1999).

El color del iris varía de unas especies a otras.



El color del iris también puede variar dentro de una misma especie con el sexo o con la edad.


El cuerpo ciliar produce un líquido que se secreta a la cámara posterior, de ahí pasa a la cámara anterior y se drena en el canal de Schlemm.
El cristalino es una lente deformable y transparente que se localiza entre la cámara anterior del globo ocular, en cuyo interior se encuentra el humor acuoso y la cámara posterior, más grande, que contiene el cuerpo vítreo. Las aves tienen además, la llamada almohadilla anular que rodea al cuerpo del cristalino.
El cristalino también tiene poder de refracción ya que su índice de refracción difiere del de los fluidos que lo rodean. Por lo tanto, los medios de refracción del globo ocular son la córnea, el humor acuoso, el cristalino y el cuerpo vítreo. Son medios transparentes que dejan pasar la luz y se encargan de proyectar la imagen en la retina.
La acomodación permite enfocar objetos que se encuentran tanto lejos como cerca. En las aves, la córnea y el cristalino intervienen en la acomodación. La contracción del músculo de Crampton varía la curvatura de la córnea (acomodación corneal) y la contracción del músculo de Brücke, empuja el cuerpo ciliar contra el cristalino provocando su deformación (acomodación lenticular).
La retina es la capa nerviosa y la que ocupa la posición más interna del globo ocular. Es más gruesa en el centro que en los extremos. La retina de las aves, a diferencia de la humana, carece de vasos sanguíneos y se nutre a partir de la coroides y del pecten.
No se puede hablar de la estructura de la retina sin nombrar al médico español Santiago Ramón y Cajal.

Antes de Cajal, se creía que las células eran individuales salvo las neuronas que se fusionaban formando una red difusa y, además, los conocimientos sobre la estructura de la retina eran escasos y confusos. Cajal demostró que las neuronas son también células individuales que se conectan por lo que el llamó «dispositivo de engranaje» y más tarde, el médico británico Charles Scott Sherrington le dio el nombre de sinapsis.
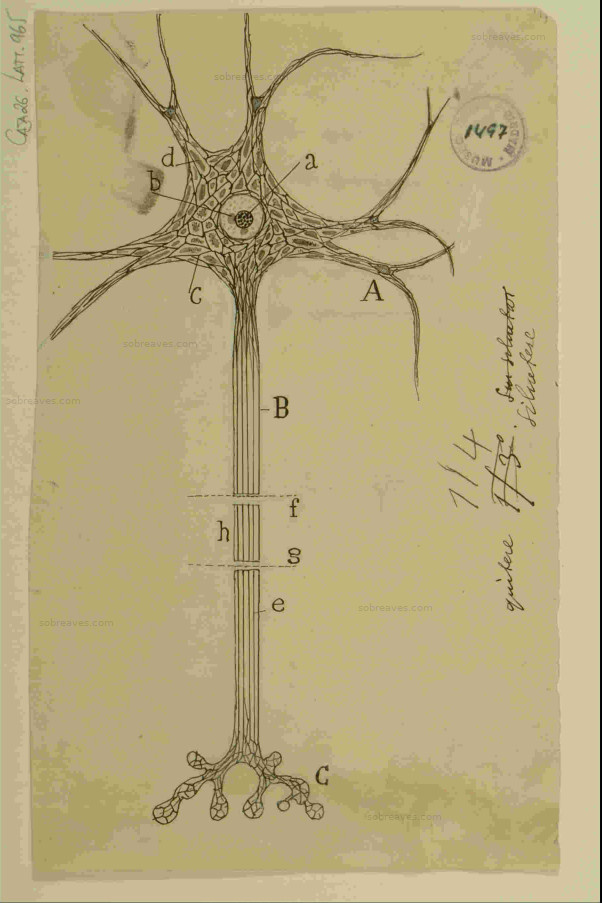
En 1893, publicó en la revista La Cellule, un artículo titulado «La rétine des Vertébrés» en donde hace una descripción de la retina tal que, aunque en los años transcurridos desde entonces, otros científicos han hecho contribuciones al estudio de la retina, su trabajo sigue vigente.
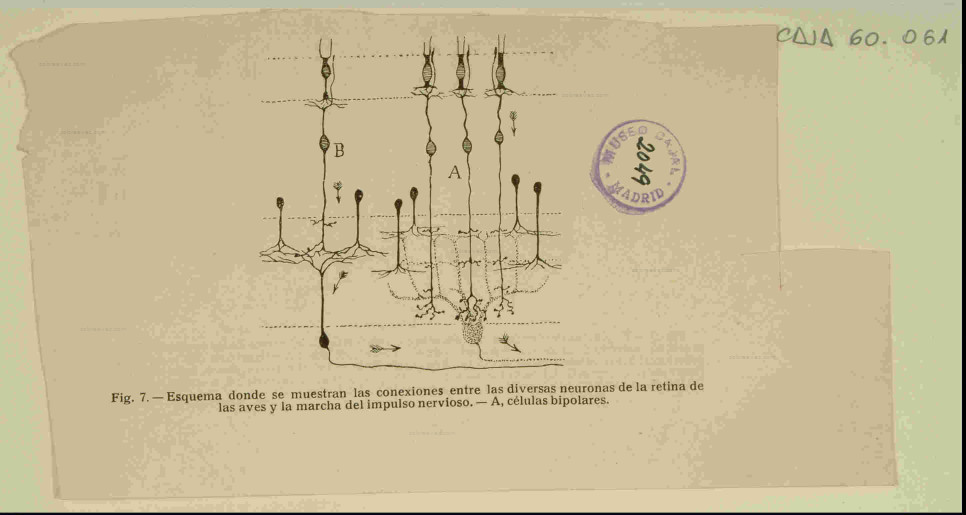
El fisiólogo italiano Marco Piccolino publicó en 1988 un artículo con el título «Cajal and the retina: a 100 years retrospective» en la revista Trends in Neurosciences, donde dice: «Hace un siglo Cajal publicó sus primeros estudios sobre la retina mediante el método Golgi. Este trabajo representa un hito en el nacimiento del campo de la neurociencia», y añade: «Para enfatizar el impacto del trabajo de Cajal basta decir que su artículo de 1893 sigue siendo un punto de partida para cualquier estudio anatómico de los circuitos de la retina».

La retina consta de un epitelio pigmentario y una parte sensorial formada, a su vez, por neuronas especializadas y células gliales.
Entre las neuronas especializadas están en primer lugar, los fotorreceptores que se encargan de transformar la energía luminosa en impulsos nerviosos, estos se transmiten a las células bipolares y a las células ganglionares y, salen del globo ocular por el nervio óptico hasta el encéfalo. Otras neuronas son las células horizontales y las amacrinas que establecen importantes conexiones sinápticas.
En cuanto a las células gliales, hay que destacar las células de Müller que realizan diferentes funciones como nutrición, sostén, mantenimiento del medio extracelular etc.
Las células retinianas se disponen en capas bien definidas:
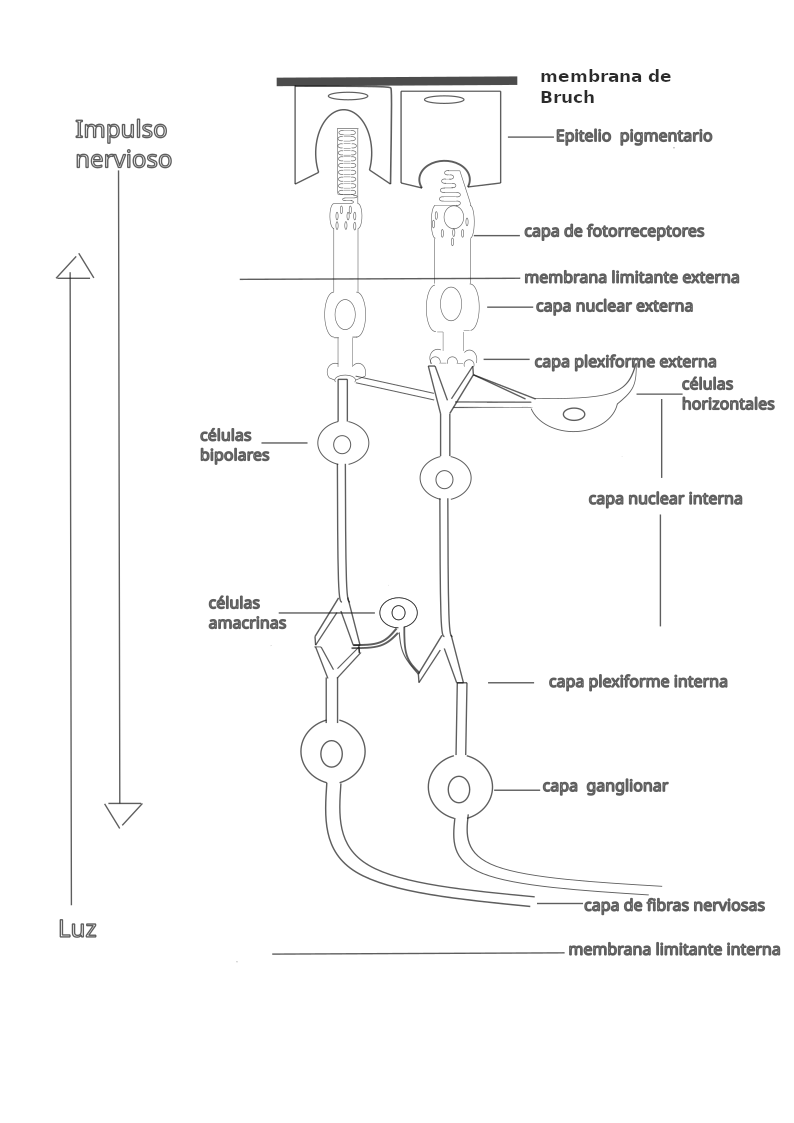
- Membrana de Bruch (BM). También llamada lamina basalis choroideae, separa la coroides de la retina.
- Epitelio pigmentario (RPE). Se sitúa cerca de la coroides. Está formado por una sola capa de células cúbicas con pigmento y, emiten prolongaciones hacia los segmentos externos de los fotorreceptores.
- Capa de fotorreceptores (IS/OS), Son de dos tipos: conos y bastones.
- Membrana limitante externa (ELM). Se localiza en la base de los fotorreceptores.
- Capa nuclear externa (ONL). En ella se encuentran los núcleos de los fotorreceptores.
- Capa plexiforme externa (OPL). Zona de sinapsis entre fotorreceptores y células bipolares y, entre células horizontales que se comunican con fotorreceptores y con células bipolares.
- Capa nuclear interna (INL). Capa donde se sitúan los núcleos de las células horizontales, bipolares y amacrinas.
- Capa plexiforme interna (IPL). Zona de sinapsis entre células bipolares, ganglionares y amacrinas.
- Capa ganglionar (GCL). Estrato que contiene los cuerpos de las células ganglionares.
- Capa de fibras nerviosas (RNFL) . Los axones de las células ganglionares forman el nervio óptico.
- Membrana limitante interna (ILM). Marca la separación entre la retina y el humor vítreo.
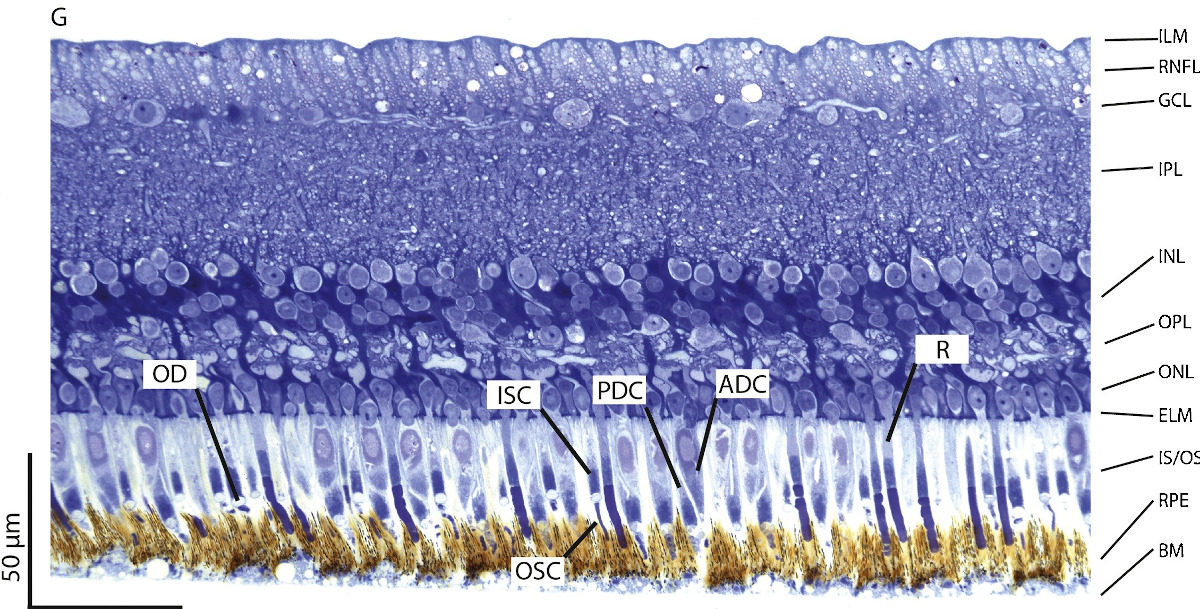
Como los fotorreceptores se encuentran en la zona más profunda de la retina sensorial, la luz tiene que atravesar las otras capas antes de llegar a ellos.
La distribución de las células en la retina no es uniforme. Existen zonas con mayor densidad de células ganglionares y fotorreceptores por lo que la agudeza visual es mayor. Dichas zonas, se llaman áreas cuando tienen forma redondeada y líneas si son alargadas. Las áreas y líneas pueden contener unas depresiones llamadas fóveas, donde la agudeza visual aún es mayor. En las fóveas abundan los conos sobre los bastones y en algunas aves, solo contienen conos.
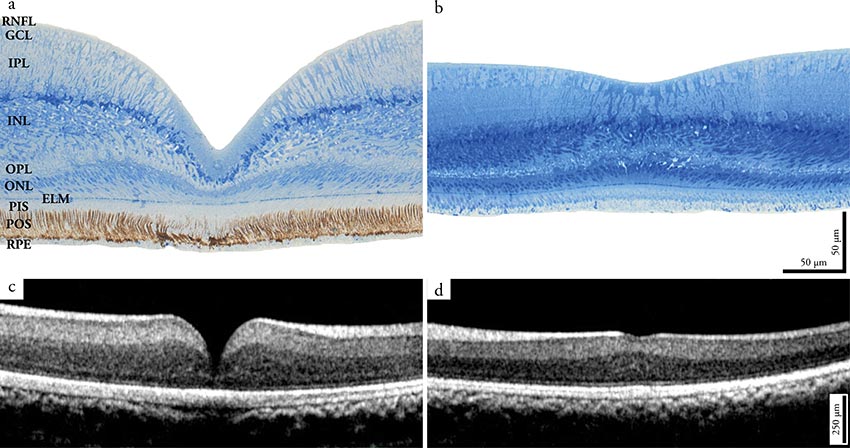
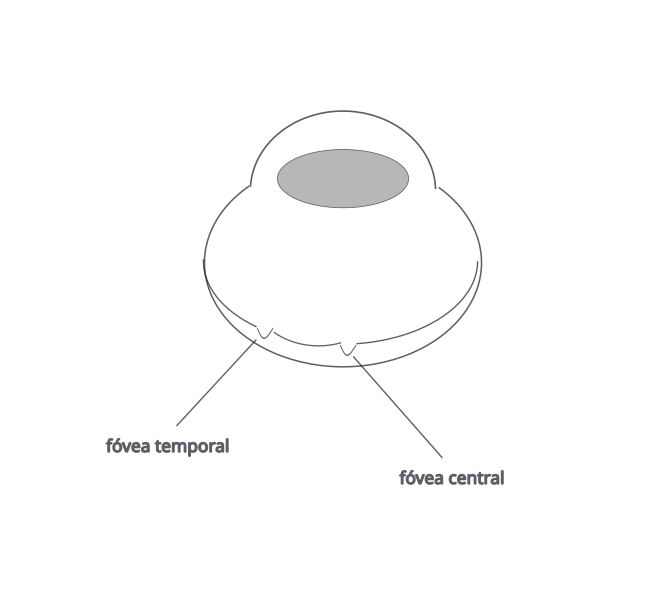
La fóvea se produce por desplazamiento de las capas superficiales de la retina para que la luz llegue mejor a los fotorreceptores. La mayoría de las aves tienen una sola fóvea que suele ser la fóvea central. Hay aves que tienen dos fóveas, una fóvea central y otra temporal. La fóvea central es más profunda y de mayor agudeza visual que la fóvea temporal. En la fóvea central se proyectan las imágenes que proceden de la visión monocular (campo de visión lateral), mientras que en la visión binocular, la imagen se proyecta en la fóvea temporal (campo de visión frontal). Aves que tienen dos fóveas son por ejemplo, rapaces (águilas, halcones…), algunos paseriformes (golondrinas…), charranes, martines pescadores…
Como muchas aves tienen muy buena vista, presentan un número mayor de células ganglionares y fotorreceptores que los demás vertebrados y las conexiones entre sus células son mucho más abundantes y complejas.
La agudeza más alta registrada hasta ahora en cualquier animal es la del águila audaz (Aquila audax) y la del buitre de pico largo (Gyps indicus). Es el doble de la agudeza más alta registrada en humanos jóvenes (Martin G.R., 2021).
Debido a que la densidad de los fotorreceptores o de las células ganglionares varían a lo largo de la retina. Los expertos elaboran mapas de contorno de isodensidad. En dichos mapas se enlazan las zonas de la retina que tienen las mismas densidades celulares (x 1000, por milímetro cuadrado). Los mapas de contorno de isodensidad, proporcionan una visión rápida del aspecto de la retina de una especie, localizan las zonas donde la agudeza visual es máxima y también permiten compararla, de manera fácil, con la retina de otras especies.
Los fotorreceptores son células especializadas que contienen fotopigmentos que absorben la luz de diferentes longitudes de onda, lo que desencadena una serie de reacciones, provocando una corriente nerviosa que viaja a los centros visuales del encéfalo, donde la sensación se hace consciente.
Hay dos tipos de fotorreceptores: conos y bastones.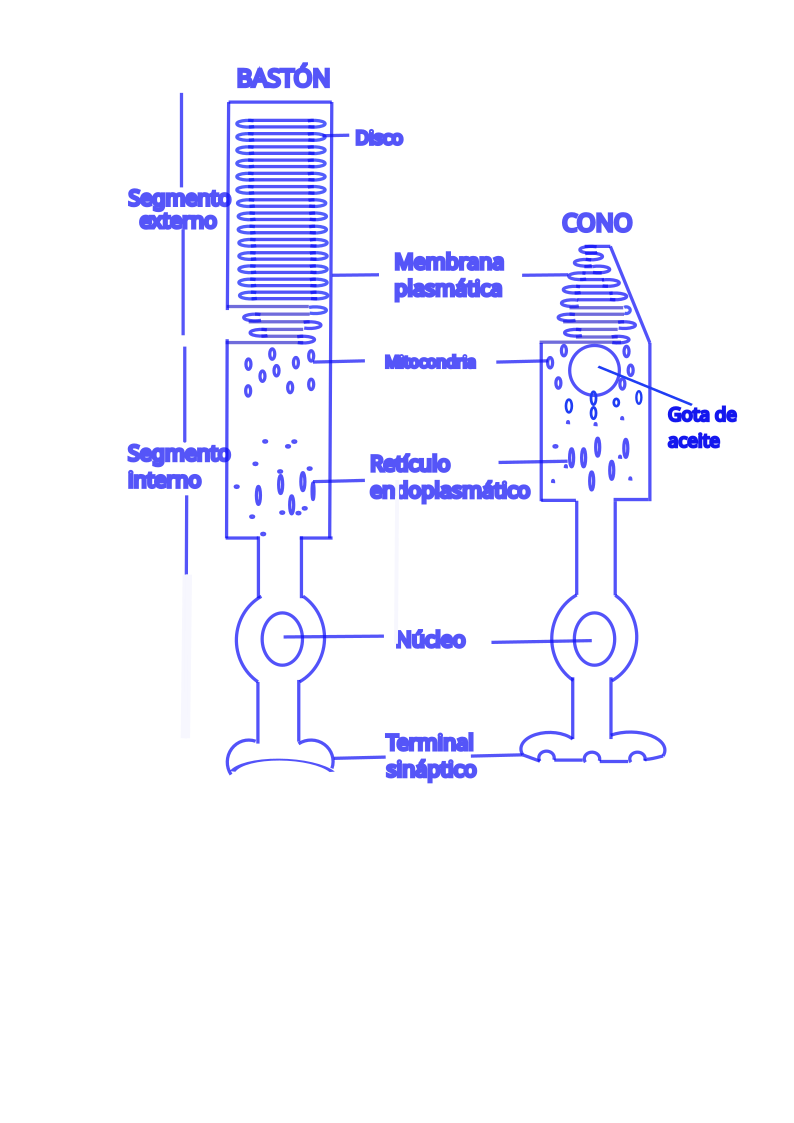
La membrana plasmática del segmento externo está muy replegada y contiene millones de moléculas de un mismo fotopigmento. Dentro de los segmentos internos, las mitocondrias se encuentran a distancia para formar el llamado elipsoide, mientras que la mayoría de los otros orgánulos, como el aparato de Golgi y el retículo endoplasmático, residen en la región proximal o mioide.
Los conos de las aves y de otros vertebrados no humanos, tienen cerca del segmento externo, una gota de aceite que puede estar coloreada por carotenoides que ingieren en la dieta. La luz pasa a través de la gota de aceite antes de llegar a la molécula de fotopigmento. La gota de aceite actúa como filtro de corte, absorbe la luz de longitudes de onda cortas y deja pasar hacia el segmento externo la luz de longitudes de onda más largas y, permite a las aves distinguir más colores.
Los fotorreceptores son bastante similares en las distintas especies de aves. Las diferencias de visión están más relacionadas con la distribución de fotorreceptores en la retina y su densidad. También hay que tener en cuenta que los fotorreceptores en las aves diurnas son estrechos favoreciendo la agudeza visual, mientras que en las aves nocturnas son más amplios, favoreciendo la sensibilidad a la luz.
Los bastones son muy sensibles a la luz, es decir, se pueden estimular con niveles bajos de luz y proporcionan una visión en blanco y negro. Abundan en la retina de las aves nocturnas. Contienen el fotopigmento RH1 con un máximo de sensibilidad espectral alrededor de los 500 nm.
Los conos necesitan para estimularse niveles más altos de luz, pero proporcionan una visión más aguda y en color, Abundan en la retina de las aves diurnas.
Los conos pueden ser simples y dobles.
Las aves tienen cuatro tipos de conos simples, es decir, son tetracromáticas, por lo que pueden ver más colores que los humanos que somos tricromáticos.
Cada cono aviar contiene un solo tipo de pigmento y un solo tipo de gota de aceite, por lo que los cuatro conos simples de las aves son:
- LWS = Cono sensible a la longitud de onda larga, contiene el pigmento m/lws y la gota de aceite es roja.
- MWS = Cono sensible a la longitud de onda media, con el pigmento rh2 y gota de aceite amarilla.
- SWS = Cono sensible a la longitud de onda corta, pigmento sws2, con la gota de aceite incolora.
- UVS/VS = Contienen el pigmento sws1 y la gota de aceite es transparente. Las aves con conos UVS tienen verdadera visión ultravioleta (UV). Son gaviotas (Laridae, Charadriiformes), avestruces (Struthioniformes), loros (Psittaciformes)) y paseriformes oscinos, excepto córvidos (Odeen y col., 2010, 2011; Odeen y Hastad, 2013). Su sensibilidad espectral máxima es menor de 400 nm.

Curruca cabecinegra/Sardinian Warbler/Curruca melanocephala Las aves con conos VS no poseen verdadera visión ultravioleta porque tienen estructuras oculares que la absorben y la luz no llega a la retina.

En 1995, Viitala y colaboradores, llegaron a la conclusión de que los cernícalos localizaban a pequeños roedores de campo, gracias a que podían detectar el reflejo UV de su orina. Esta creencia se extendió a otras rapaces.

En 2013, Lind y col., publicaron un artículo en la revista Journal of Experimental Biology, titulado «Ultraviolet sensitivity and colour vision in raptor foraging» A estos autores les parecía una contradicción que las rapaces pudiesen ver el reflejo UV de la orina de pequeños roedores y no pudiesen ver el reflejo UV de las plumas de pájaros cantores, como los herrerillos que utilizan dicho reflejo. como señales de comunicación «ocultas» a sus depredadores, sobre todo en épocas de reproducción. Estudiaron cuatro especies de rapaces: ratoneros comunes (Buteo buteo), gavilanes (Accipiter nisus), milanos reales (Milvus milvus) y cernícalos (Falco tinnunculus).

No encontraron evidencias de que estas rapaces pudiesen ver el reflejo UV de la orina de ratones de campo. Por otro lado, la visión UV disminuye la agudeza visual debido a lo que los expertos conocen como aberración cromática. Teniendo en cuenta que entre las rapaces diurnas se encuentran aves que tienen la mayor agudeza visual conocida, los científicos buscaron algún mecanismo que minimizara la aberración cromática, no lo encontraron. En la actualidad se sabe que las rapaces tienen conos VS, no pueden ver la luz UV porque sus córneas y cristalinos la absorben y no llega a la retina.
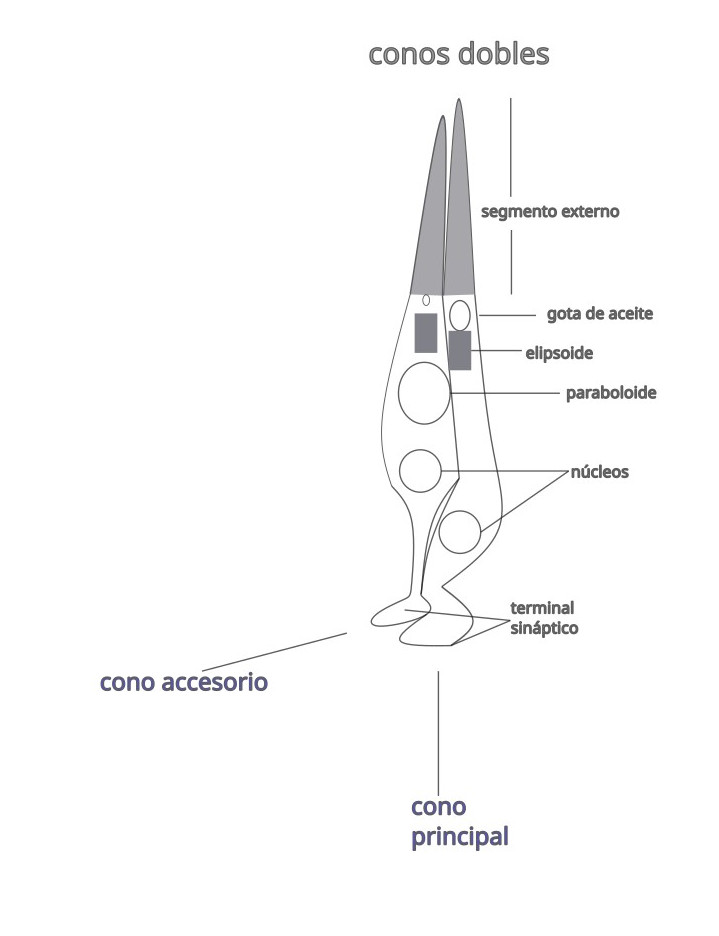
Los conos dobles se encuentran en la mayoría de los vertebrados excepto en mamíferos placentarios. Constan de dos conos acoplados: el cono principal más alargado y el cono accesorio más corto. En el cono principal hay una depresión donde se aloja parte del cono accesorio. Ambas células contienen el mismo fotopigmento (lw) con un máximo de sensibilidad espectral a 565 nm. Los conos dobles no intervienen en la visión en color, se cree que participan en la detección del movimiento.
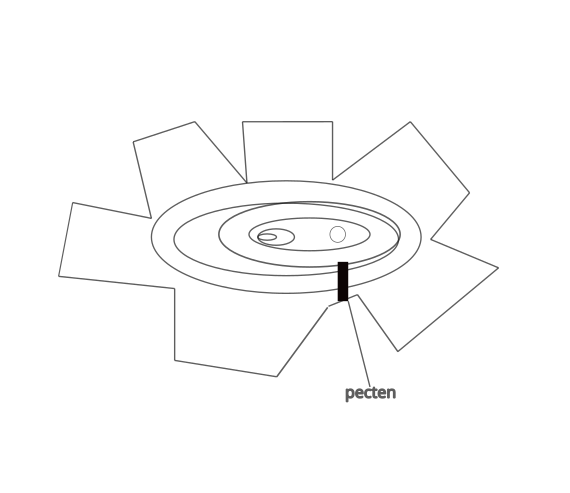
Los mamíferos tenemos vasos sanguíneos en la retina para nutrir las células nerviosas, salvo los fotorreceptores, de cuya nutrición se encargan los vasos sanguíneos de la coroides. La retina de las aves es avascular, pero posee una estructura llamada pecten (peine) oculi, cuyo descubrimiento se atribuye al francés Claude Perrault (1676). El pecten es rico en vasos sanguíneos y se encarga de nutrir a las células retinianas. También, se le han atribuido otras funciones, una de las que cuenta con mayor apoyo experimental es la regulación del pH intraocular.
El pecten está rodeado por la membrana vitreopectineal que se considera una continuidad de la membrana limitante interna de la retina.
El pecten es una estructura que está muy plegada, esto le permite ocupar poco espacio y, junto con su ubicación próximo al nervio óptico, hace que interfiera menos en la visión que la circulación retiniana de mamíferos. El plegamiento, a su vez, le proporciona una gran superficie de intercambio, para que difundan nutrientes y oxígeno a las células y recoger los desechos del metabolismo.
El número de pliegues se asocia con la actividad del ave y sus necesidades. Así, en las aves diurnas el pecten tiene más pliegues que en las aves de actividad nocturna. De las aves estudiadas hasta ahora, el mayor número de pliegues se encontró en especies del género Corvus, como el cuervo americano y, el menor número en Caprimulgus como el chotacabras.
El pecten es de color oscuro, más en el ápice que en la base, debido a la presencia de células pigmentadas, los melanocitos. En la superficie del pecten se han identificado células con actividad fagocítica, los hialocitos, que se considera que son un subtipo de macrófagos.
Por su forma se distinguen tres tipos de pecten:
- Pecten oculi plicatus = Es la forma más estudiada ya que se encuentra en la mayoría de las aves. Consta de una base, los pliegues dispuestos en acordeón o abanico invertido y el puente, muy pigmentado, que une los pliegues.
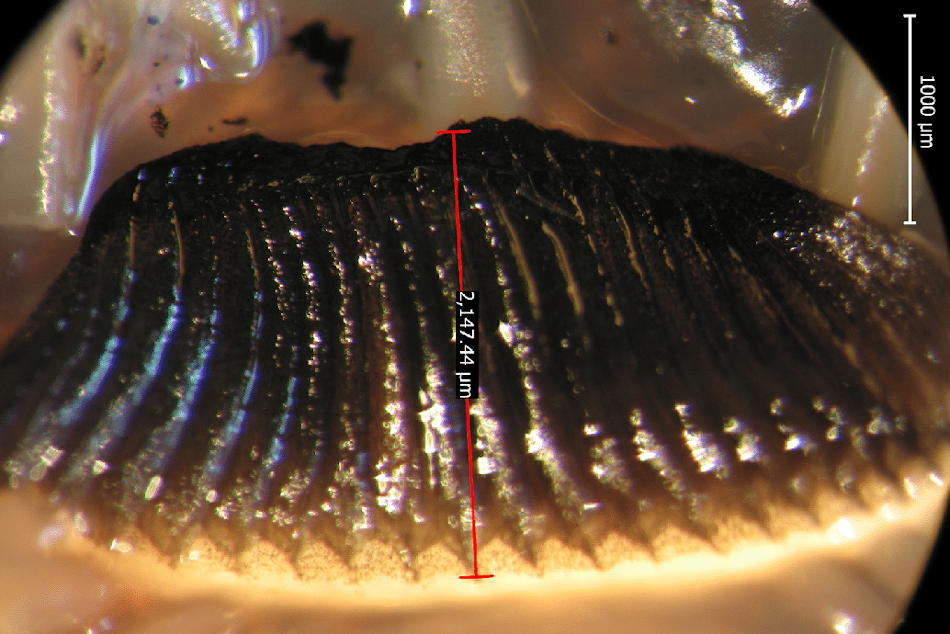

- Pecten oculi vanellus = Típico del avestruz, ñandú y tinamú. Tiene una ramificación bipennada. Kiama y col. (2006), lo describen formado por una laminilla primaria vertical gruesa de la que parten laminillas secundarias más delgadas, dispuestas a ambos lados. Las laminillas secundarias, que van desde la base, confluyen en el ápice dándole aspecto de cono. De algunas laminillas secundarias parten laminillas terciarias. La pigmentación es mayor en los extremos distales de las laminillas secundarias y terciarias.
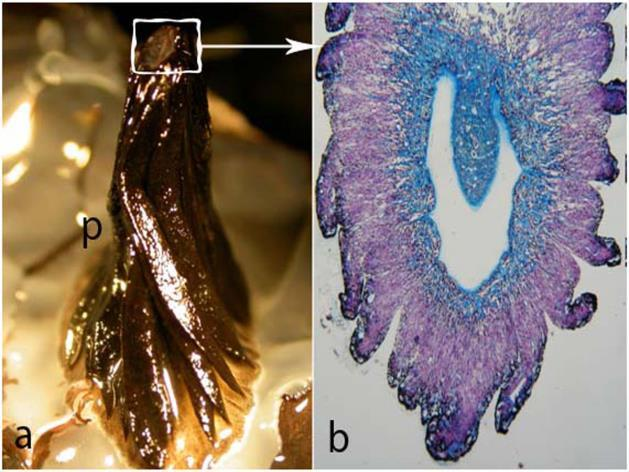
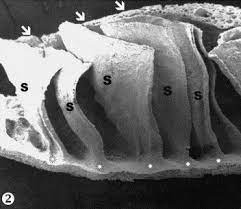
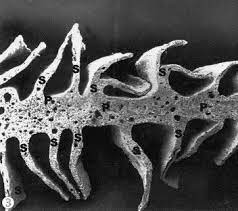
- Pecten oculi conicus = Presente en el kiwi. Para Walls (1942), se trata de un cono delgado, sin pliegues ni laminillas, muy parecido al corpus papillaris de muchos lagartos.
Excepcionalmente, algunas aves con hábitos crepusculares o nocturnos, como los chotacabras, pueden mejorar su visión en condiciones de poca luz, gracias a una estructura reflectante llamada tapetum lucidum. El tapetum lucidum actúa como un espejo, refleja la luz que atraviesa la retina haciendo que la vuelva a atravesar. El tapetum lucidum está presente también en mamíferos, como en los gatos, y es la causa por la que al reflejar la luz, le brillan los ojos en la oscuridad.

Con permiso de María
Con permiso de María

Con permiso de María
Campos visuales
Graham Martin es uno de los mayores especialistas en campos visuales de las aves por lo que parte del texto que se expone a continuación, se extrajo de sus publicaciones.
Los campos visuales son las regiones del espacio, alrededor de la cabeza, de las que se pueden recuperar información visual en cualquier instante.
En los campos visuales se distinguen varias regiones:
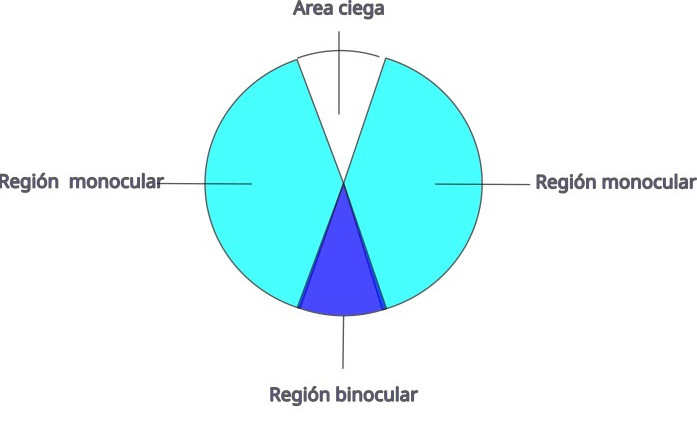
- Dos regiones laterales (monoculares). Son las de mayor agudeza visual ya que pueden abarcar la fóvea. En el caso de aves que tienen dos fóveas, correspondería a la fóvea central. La región lateral monocular se utiliza para la detección de alimento y depredadores, también interviene en el control del vuelo.
- La región frontal de superposición binocular. En nuestra especie, los ojos están dispuestos frontalmente, sus ejes ópticos son paralelos, por lo que gran parte de la imagen que capta un ojo lo capta el otro simultáneamente. Hay, por lo tanto, superposición binocular que el cerebro interpreta como estereopsis (visión en 3D). Todas las aves tienen los ojos colocados lateralmente, incluidas las rapaces nocturnas, por lo que los ejes ópticos divergen. Sin embargo, en la visión frontal, que se corresponde con la periferia del campo visual de cada ojo, hay superposición binocular. En la actualidad, la estereopsis solo se ha documentado en algunas especies nocturnas como la lechuza (Tyto alba).
Las aves necesitan manejar con exactitud el pico o las patas para conseguir alimento, construir nidos complejos y alimentar a los pollos. Esto se logra gracias al campo de flujo óptico en la región binocular. Los campos de flujo óptico describen la manera en la que la imagen del mundo se mueve a través de la retina mientras la cabeza se mueve a través del espacio. El campo de flujo óptico se expande simétricamente sobre el punto en el que el observador se está moviendo, por lo tanto, especifica la dirección del viaje y el tiempo para contactar con un objetivo. Nosotros lo podemos experimentar cuando vamos en coche o corremos y, nos parece que los objetos cercanos se mueven con más rapidez que los que se encuentran más alejados.
Cuando las aves quieren localizar un objetivo, como alimento o depredadores, utilizan la visión lateral monocular de mayor agudeza visual que suele depender de la fóvea central, pero cuando quieren alcanzar el objetivo, cambian a la región binocular frontal que, aunque de menor agudeza visual, le permite llevar el pico o las patas con precisión hasta dicho objetivo. En las aves que tienen dos fóveas, la visión binocular puede alcanzar la fóvea temporal.
- Áreas ciegas. En los campos visuales también se pueden encontrar áreas ciegas, localizadas por encima y por detrás de la cabeza, así como un área ciega anterior, que se sitúa detrás de la región binocular. Tyrrel y Fedez-Juricic han estudiado esta área ciega que se extiende desde los ojos hasta el inicio del campo binocular. Existe una relación entre el ancho del campo binocular y la longitud del área ciega anterior, es decir, cuanto más ancho es el campo binocular, más corta es dicha área ciega y más cerca está el pico del origen del campo binocular. Por lo tanto, a medida que se acorta el área ciega anterior, las aves tienen un mayor control visual del pico. Según estos autores, a medida que el campo binocular se amplia y la longitud del área ciega anterior se acorta, el área ciega detrás de la cabeza se ensancha, lo que reduce la cobertura visual general.
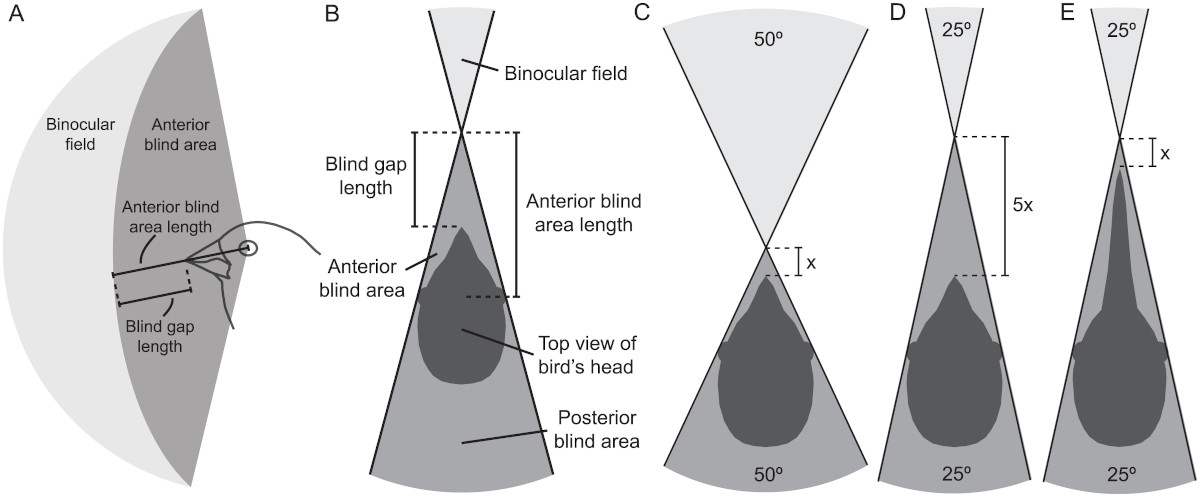
Las aves que tienen ojos grandes, están más expuestas al deslumbramiento por el sol que las de ojos pequeños y, para protegerse, cuentan con amplias áreas ciegas sobre la cabeza y anexos ópticos, por ejemplo, águilas y avestruces tienen cejas extensas que les dan sombra a los ojos y las avestruces tienen, además, pestañas (filoplumas) largas y gruesas. Los buitres, como el buitre leonado, poseen crestas de piel a modo de cejas.


Cuando aves como por ejemplo, águilas o buitres, vuelan por espacios abiertos buscando alimento, inclinan la cabeza hacia abajo y, el área ciega sobre la cabeza les impide ver lo que está delante en la dirección del viaje, esta es la causa de la muerte de muchas de ellas al colisionar con construcciones humanas como postes eléctricos y aerogeneradores. Así, Gerardo Báguena en un artículo publicado en la revista Quercus en noviembre de 2023, contabiliza 8.960 aves muertas por impacto con las aspas de aerogeneradores, en los tres últimos años, utilizando los datos oficiales de doce comunidades autónomas. Hay que tener en cuenta que no todos los cadáveres son detectados porque desaparecen a las pocas horas por degradación o depredación. Báguena también dice lo siguiente: «Mientras tanto, ya existen medidas preventivas eficaces que deberían ser exigidas por las administraciones de forma obligatoria a las empresas promotoras; todas ellas están al alcance de los consorcios que se dedican al negocio eólico. Sin embargo, en la actualidad, la normativa no exige esas medidas sino otras mucho más laxas que de todos es sabido que no están funcionando bien» Gerardo Báguena es director de la Fundación para la Conservación del Quebrantahuesos (FCG).
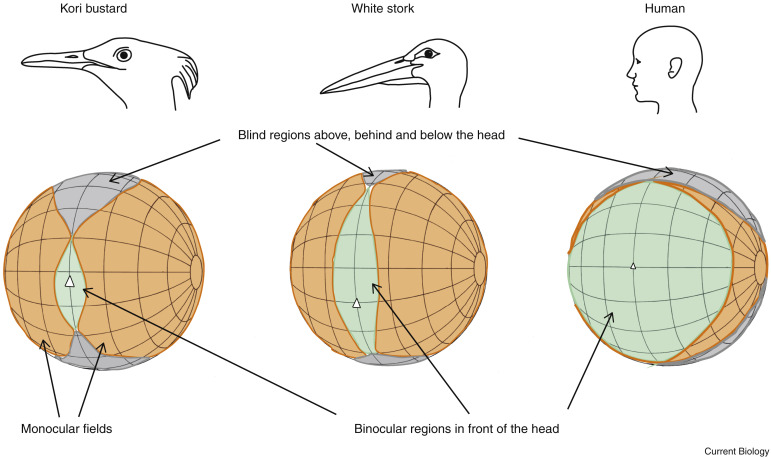
Todas las aves tienen los ojos dispuestos lateralmente pero, las posiciones en el cráneo varían entre especies, esto da lugar a una gran diversidad de configuraciones de campos visuales.
La mayoría de las especies no paserinos, tienen campos binoculares con una anchura máxima en el plano horizontal entre 20º y 30º.

La región binocular en el plano vertical es más extensa, varía entre los 60º (por ej. avutardas) y los 180º de las garzas. En el caso de las garzas, el pico se encuentra aproximadamente en el centro del campo binocular. Cuando están de pie con el pico horizontal y la cabeza ligeramente adelantada pueden ver lo que pasa a sus pies.


Mientras que hay aves que no pueden ver la punta de su propio pico, hay otras que pueden examinar lo que está entre sus mandíbulas, como es el caso de cálaos, rayadores o estorninos.

Estos campos visuales constan también de un área ciega detrás de la cabeza, cuya anchura en el plano horizontal varía de unas especies a otras. Las aves que presentan estas configuraciones de campos visuales , pueden adquirir el alimento de diferentes maneras, por ejemplo, la paloma bravía consigue alimentos inmóviles mediante picoteo, otras, como las garzas, atrapan a las presas con el pico o, con las patas como las águilas y, excepcionalmente, por filtración, como es el caso de los flamencos.
Otro tipo de campos visuales lo presentan aquellas aves que no necesitan el control visual de la posición del pico ya que se guían por señales táctiles. Sus ojos están situados ligeramente hacia arriba (alto del cráneo). Es el caso de patos (Anatidae) con picos filtradores como el pato azulón y de aves playeras (Scolopacidae), como la chocha perdiz (Scolopax rusticola) que busca comida en el fango confiando en señales táctiles y de sabor.

Sus nidos son sencillos y sus pollos son precoces, es decir, se alimentan por sí mismos poco después de la eclosión.

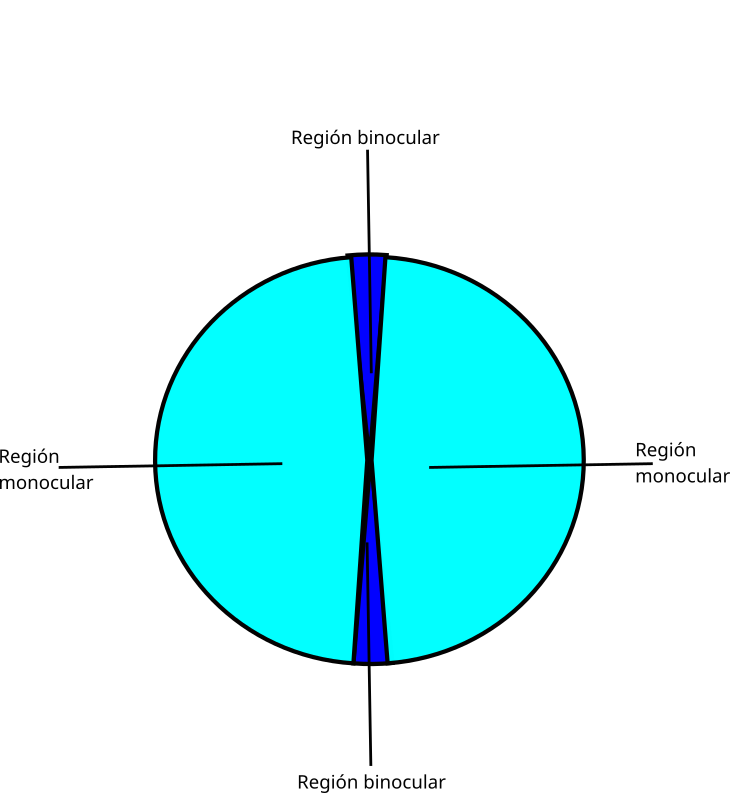
Su campo binocular en el plano horizontal es estrecho, entre 5º y 10º, lo que no les impide volar rápido incluso en bosques y con poca luz.
Tienen visión panorámica, sin áreas ciegas ya que el campo binocular se extiende 180º por delante, por encima y por detrás de la cabeza, pero la proyección del pico cae fuera o en la periferia del campo visual. El contar con una cobertura visual completa del hemisferio celeste es importante para la detección de depredadores.

Los campos visuales pueden diferir entre especies próximas, como ocurre con el pato cuchara y el silbón europeo. Mientras que el primero se alimenta por filtración y cuenta con visión panorámica, el silbón europeo necesita el control visual del pico ya que se alimenta picoteando, el campo binocular es más ancho que el del pato cuchara y tiene un área ciega por detrás de la cabeza.
Los flamencos, aunque se alimentan por filtración, carecen de visión panorámica. Su campo binocular abarca la punta del pico que utiliza con gran precisión para construir nidos complejos y alimentar a sus pollos con leche de buche. Tienen también un área ciega detrás de la cabeza.

El correlimos gordo, al igual que la chocha perdiz, utiliza señales táctiles desde la punta de su pico para alimentarse en sus zonas de invernada. No tiene visión panorámica porque en sus áreas de reproducción se alimentan de invertebrados voladores o de superficie, que requiere un control visual del pico. Tienen un pequeño campo binocular alrededor del pico y un pequeño sector ciego detrás de la cabeza.

Los campos binoculares más anchos en el plano horizontal, entre 40º y 50º, se encuentran en rapaces nocturnas y paseriformes. El campo binocular más ancho lo tiene el cuervo de Nueva Caledonia (60º) ya que fabrica y utiliza herramientas para extraer larvas del interior de troncos.
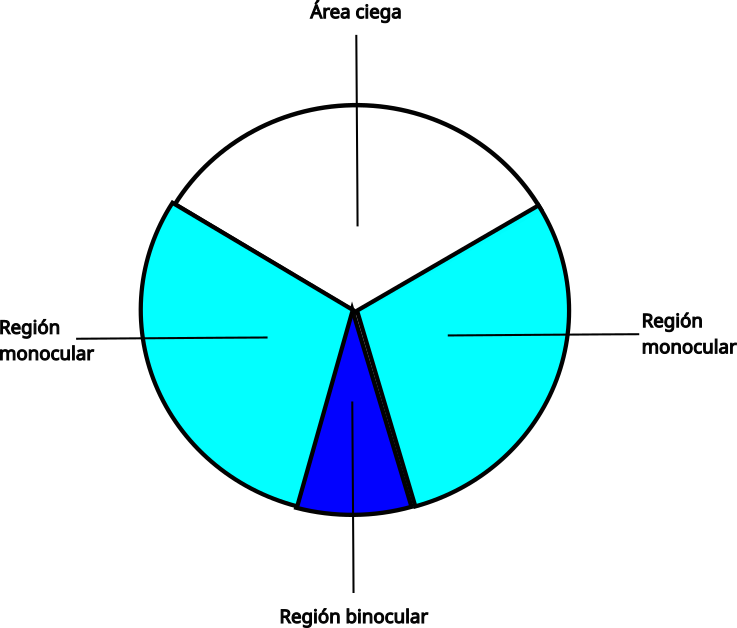 El área ciega detrás de la cabeza es muy amplia en las rapaces nocturnas, se cree que se debe a las elaboradas estructuras del oído externo que impiden una posición más lateral de los ojos.
El área ciega detrás de la cabeza es muy amplia en las rapaces nocturnas, se cree que se debe a las elaboradas estructuras del oído externo que impiden una posición más lateral de los ojos.

Phalacrocorax carbo
La configuración de los campos visuales varía con el movimiento de los ojos. A diferencia de los humanos, las aves no mueven los ojos de manera coordinada, mientras que dirigen un ojo hacia delante, el otro lo pueden mover hacia atrás. Cuando los dos ojos miran al frente, la amplitud del campo binocular en el plano horizontal aumenta. Sin embargo, cuando los dos ojos miran hacia atrás, la anchura del campo binocular disminuye e incluso, puede llegar a desaparecer. También pueden variar las dimensiones del área ciega. En el caso de aves como por ejemplo, el cormorán grande, además del movimiento de traslación de los ojos, también lo tiene de rotación, por lo que puede llegar a hacer desaparecer la superposición binocular, pero sin variar apenas la amplitud del área ciega.
No todas las aves pueden mover los ojos como es el caso, por ejemplo, de las rapaces nocturnas. Esto lo suplen con una gran flexibilidad en el cuello que pueden girar hasta unos 270º.
Para facilitar su estudio, los científicos representan los campos visuales como una esfera con un sistema de coordenadas angulares basado en la latitud y longitud convencionales y, en cuyo centro se encuentra la cabeza del ave.
Sueño unihemisférico
Hay aves, al igual que m amíferos marinos, que pueden dormir con un ojo abierto, es decir, duermen con la mitad del cerebro mientras que la otra mitad está despierta. Esto se conoce como sueño de ondas lentas unihemisférico. El sueño unihemisférico no les impide a las aves realizar bien tareas como estar alerta ante la presencia de depredadores, comer o, en aves que realizan largas migraciones sin escalas, poder dormir mientras vuelan. Rattenborg (1999) observó que, cuando un grupo de patos azulones se encontraba posado, los que ocupaban el centro utilizaban mucho menos el sueño unihemisférico que los que se situaban en la periferia, ya que esos últimos se mantenían vigilando por si se acercaba un depredador.
amíferos marinos, que pueden dormir con un ojo abierto, es decir, duermen con la mitad del cerebro mientras que la otra mitad está despierta. Esto se conoce como sueño de ondas lentas unihemisférico. El sueño unihemisférico no les impide a las aves realizar bien tareas como estar alerta ante la presencia de depredadores, comer o, en aves que realizan largas migraciones sin escalas, poder dormir mientras vuelan. Rattenborg (1999) observó que, cuando un grupo de patos azulones se encontraba posado, los que ocupaban el centro utilizaban mucho menos el sueño unihemisférico que los que se situaban en la periferia, ya que esos últimos se mantenían vigilando por si se acercaba un depredador.